



When Life Doesn’t Give You Neoblasts, Regenerate Anyways! (Fixations #1)
In the first of a new series, we take a dive into the strange stem cell system of the catenulid flatworm Stenostomum brevipharyngium.

Background
When you think of regeneration, the first examples that come to mind are usually of a lizard losing its tail or an axolotl regrowing a limb. While these organisms are definitely hallmarks for the study of regeneration, there is a wide diversity of regenerative capability across the animal kingdom. In fact, some of the most regenerative organisms are probably ones you don’t think of: worms! Flatworms, such as planarians, have become a popular model for the study of regeneration due to their ability to relentlessly regrow their heads and tails, even if they’re cut into multiple pieces. They can do this thanks to neoblasts, a population of adult pluripotent stem cells that can help rebuild tissue that is lost. These stem cells are unique in that they can differentiate into any cell type in the worm throughout their lifespan. They are also characterized by something called the germline multipotency program, a group of canonical markers expressed in these stem cells that help maintain their stemness, or their identity as stem cells. Adult pluripotent stem cells aren’t just unique to flatworms, either–they’ve been found in organisms as diverse as cnidarians and sponges.
However, while adult stem cells have been identified in a battery of organisms with significant regenerative capabilities, how they came about evolutionarily still remains a mystery. Did the ancestor to all of these organisms possess a similar stem cell population, or did these unique stem cells evolve independently across the animal kingdom on multiple occasions? To begin to get at the heart of this question, members of Max Planck Institute’s Rink Lab and Stanford University’s Wang Lab decided to study the stem cells of Stenostomum brevipharyngium, a member of a group of flatworms called Catenulida. Previous studies characterizing these stem cells have only focused on their morphology, meaning that how they contribute to regeneration and the characteristic genes they express remain unknown. By studying these two aspects of these stem cells, then, the authors hoped to better understand the evolutionary context behind them (i.e. if they express the same genes as adult pluripotent stem cells in other species) and if their function is conserved (i.e. if they contribute to regeneration in similar ways).
Main Findings
The authors began by characterizing regeneration in Stenostomum following an amputation. They found that, like other highly regenerative organisms, the flatworm is capable of regenerating major structures like sensory organs and musculature very rapidly—within the first four days following amputation. The authors wanted to understand, however, how cells capable of dividing were contributing to the regeneration of new tissue. To do so, they used a method commonly applied to other flatworm models called X-ray irradiation, which eliminates pluripotent stem cells by damaging their DNA and preventing cell proliferation, in turn halting regeneration and cell turnover. Stenostomum, while surprisingly resistant to irradiation, did suffer tissue regression following treatment, indicating that they rely on actively dividing cells to fuel regeneration as has been established in other flatworm models.
Having established these commonalities, the authors decided to turn towards the molecular side of things and better define the expression profiles of these cells. They began by using sequencing techniques to interrogate what genes were being upregulated or downregulated during irradiation and following irradiation treatment. Using similar datasets from other flatworm models, the authors defined a core irradiation-sensitive gene set, which featured genes involved in cellular processes like mitosis and ribosome biogenesis. Interestingly, however, the authors noticed that many of the markers that make up the germline multipotency program—genetic hallmarks of adult pluripotent stem cells—were missing from this set. This marked the first of what would be multiple intriguing diversions from what has been characterized in other models.
The authors proceeded to return to the actively dividing cell populations and characterize them further. To do so, they used two different molecular staining techniques—EdU staining and H3P immunohistochemistry—to label dividing cells throughout Stenostomum. Here, they were met with another twist: they identified several dividing cell populations in multiple different tissues of the flatworm. This was a particularly surprising finding given that pluripotent stem cells in other flatworm models are the only cells with competency for division and do not reside within differentiated tissues or organs. To dig further into this mystery, the authors decided to perform a more specific sequencing technique in order to characterize the different cell populations.
Using a sequencing technique called whole-body single-nuclei RNA sequencing, the authors were able to assemble a cell atlas that identified 18 presumptive cell clusters in Stenostomum. Of these, only one of these clusters–cluster 4–showed extreme sensitivity to radiation. Further analysis showed that these cells were localized lateral to the intestine–specifically, within the epidermis, or skin–and that the genes they express are related to cell division and ribosome synthesis. Before diving further into this population, though, the authors wanted to compare their cell atlas to a planarian dataset. Here, they found that cluster 4 bore the greatest resemblance to planarian neoblasts (or, adult stem cells); however, this cluster lacked the canonical markers of the germline multipotency program typically expressed by neoblasts. Digging further into this, the authors found that while some characteristic markers were not expressed at all, such as piwi, other components of this program, such as argonaute, were actually expressed broadly in multiple cell types.
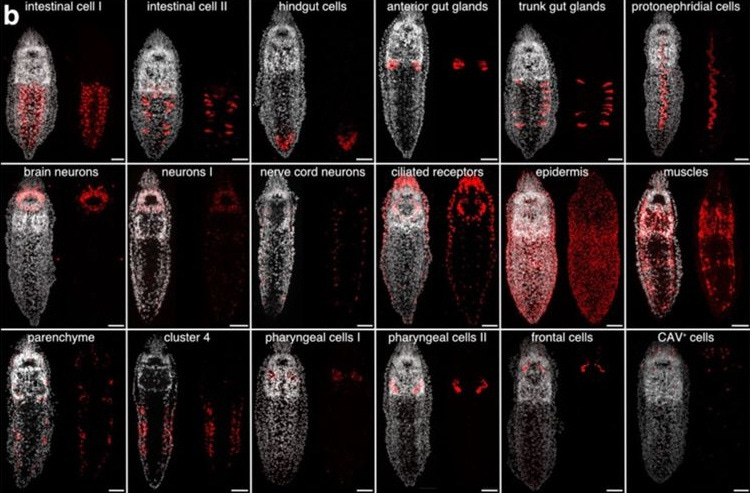
In order to better understand the dividing cell populations they identified in Stenostomum, the authors decided to examine the expression of three different markers from cluster 4 of their atlas: go_Sbre_v1_33587_1_1 (nicknamed cluster 4 marker 1 or C4M1), fibrallarin (FBL) and histone H2A (H2A). All three were co-expressed with dividing cells located along the gut and in the epidermis. However, further experiments revealed interesting patterns of co-expression or a lack thereof, leading the authors to define four distinct cell populations: “deep” stem cells that express C4M1, epidermal stem cells that co-express both stem cell and differentiated marker genes, division competent cells that completely lack stem cell markers, and organs devoid of division competent cells.
To make sure that the markers associated with the germline multipotency program really weren’t involved in Stenostomum’s stem cell system, the authors designed probes to visualize the expression of piwi, vasa, and nanos. While these experiments did identify small clusters of vasa+/piwiA+ cells in the epidermis, these cells were not actively dividing and did not express C4M1. To test whether these markers were being expressed transiently during early stages of regeneration–a phenomenon that has been previously observed in a different worm model–the authors assessed expression of the markers at multiple time points of head regeneration in Stenostomum. Though they detected cells positive for C4M1 and FBL starting at 5 hours post amputation, they did not detect any change in expression for the stem cell markers that would indicate contributions to regeneration. The marker, nanos, however, proved a somewhat more complex case. The authors had previously noticed that nanos was highly upregulated following irradiation treatments. When they assessed expression of nanos along with other germline multipotency markers, they found that nanos signal increased dramatically in cell clusters positive for piwiA. Interestingly, these clusters also became positive for their presumed stem cell marker, C4M1. Together, these findings led the authors to conclude that the cell clusters expressing germline multipotency markers may actually represent the transition of epidermal cells into transitory gonads, a phenomenon that has been shown previously in Stenostomum.
To round out their study, the authors decided to use a technique called dsRNA soaking to interfere with the RNA of their markers of interest and see if there is a functional consequence on the cell clusters. They found that knocking down piwiA or piwiB had no effect on expression of C4M1, showing that these markers are not necessary for the maintenance and function of C4M1+ division-competent cells (despite the fact that piwiB is expressed in some of these cells). They also found that these knockdowns had no effect on cell proliferation, a surprising result given that piwi has been previously established as having a role in maintenance of cell proliferation. To widen their analysis beyond these markers, the authors also decided to use cell cycle regulator cdk1 and additional germline multipotency component bruno-like to their screen. They found that knockdown of cdk1 prohibited regeneration from occurring, while knockdown of bruno-like had no effect. Overall, the authors concluded that the germline multipotency program is not required for processes like regeneration or asexual reproduction in Stenostomum and that it is not involved with the worm’s presumptive stem cell system.
The Takeaways
So, why should we be intrigued by Stenostomum’s stem cell system? As the authors have shown throughout this pre-print, these stem cells represent a major departure from the neoblasts characterized in other flatworms. Not only do these stem cells lack a canonical multipotency program characterized in neoblasts, they are also made up of multiple cell populations that reside in differentiated tissues and organs. While more analysis will be needed to fully determine the potency of these cells (i.e. the extent of their capability for differentiation) and how they contribute to regeneration, this study marks an exciting step towards understanding what has proved an already unique stem cell system.
This study also raises important questions regarding the evolution of adult pluripotent stem cells. Catenulida, an order of flatworms, is considered a sister group (or, the closest relative) to Rhabditophora, a group containing all other flatworms. If these other flatworms have neoblasts, what happened evolutionarily? There are two possibilities: catenulids like Stenostomum underwent some kind of evolutionarily loss in addition to changes in their cell types and germline multipotency program, or catenulid stem cells represent the ancestral state and other flatworms may have co-opted the germline multipotency program to give rise to their neoblasts. If it’s the former, this could mean that the stem cell systems we’ve come to know in different organisms derive from some kind of ancestral system. If it’s the latter, this could mean that stem cell systems in multiple organisms could have arisen independently several times over evolutionary history. Either way, this will provide us with more information regarding how vertebrates like us have lost our regenerative capabilities and if there are ways to reactivate them. Until then, establishing and characterizing models like Stenostomum will be important for understanding the nature of these stem cells and how they came about.